| RIKEN Center for Developmental Biology (CDB) 2-2-3 Minatojima minamimachi, Chuo-ku, Kobe 650-0047, Japan |
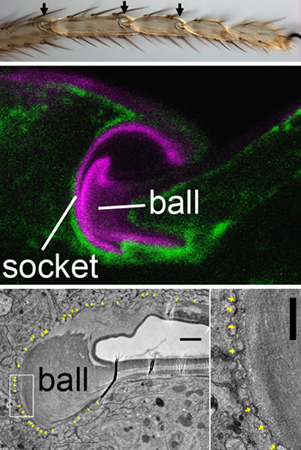
June 15, 2010 Insects wear their skeletons on the outside, but their chitinous cuticles must perform many of the same functions as vertebrate bones. In addition to their protective functions, exoskeletons also provide structural support and defined range of motion for limbs. This last role is the duty of the joints, which as in vertebrate limbs, come in various configurations and play central roles in such behaviors as mobility and feeding. (Indeed, an entire phylum of invertebrates, the Arthropoda, was originally named after its members jointed feet.) But while the physiology of vertebrate joints has been extensively studied, much less is know about their development. Reiko Tajiri in the Laboratory for Morphogenetic Signaling (Shigeo Hayashi, Group Director) in collaboration with the Electron Microscopy Laboratory (Shigenobu Yonemura, Laboratory Head) chose the leg of the fruit fly Drosophila as a model to study the formation of ball-and-socket joint skeletons. Using a combination of cell labeling and imaging technologies, the group showed that this structure forms through a sequential process involving extensive cell migration and tissue remodeling.
The study began with some basic observations, as the development of joint cuticles is largely unexplored. After characterizing the morphology and cell arrangements in the mature joint by optical and electron microscopy, Tajiri looked at the same region earlier in development to watch how these come to be. The leg primordium begins as grooved epithelium that gradually lengthens, with points of constriction at the future joints. The joints themselves form when cells on the dorsal side constrict more tightly and fold inward in a proximal direction, deepening the furrow into an empty cavity. Once this cavity has formed, the cells that line it begin to deposit cuticle, forming the ball at the deepest part of the cavity, and the socket on the dorsal side. The socket cuticle later expands ventrally toward the bottom of the cavity, and eventually covers the entire surface. The group next turned to a structural feature known as plasma membrane plaques (PMPs), which have been considered to be the sites of chitin release and of the adhesion between the cuticle and underlying cells. The question Tajiri sought to answer was, do the cuticle-forming activity move like a wave over a bed of stationary cells, or do the cells themselves move? The group used a genetic labeling method to monitor the locations of cells within the joint region at various stages, and identified three drivers of reporter gene expression in distinct, but partially overlapping, subpopulations. By tracking the behavior of the labeled cells, they determined that the apical surfaces of cells making up the ball and socket components of the joint followed the distribution of PMPs. The next step was to see which parts of the cuticle were produced by which of the genetically marked regions. Tajiri and colleagues found that one subpopulation (expressing fng) was specific to the dorsal aspect of the socket, while the other two, neur- and bib-, overlapped, with neur-expressing cells giving rise to ventral socket while bib-expressing cells formed the cuticle of the ball. In contrast to the cuticle, the surfactant that lubricates the joint seemed to be manufactured more diffusely. Perhaps unsurprisingly, blockade of chitin synthesis disrupted the entire joint cuticle. But interestingly it had no effect on the cuticle-secreting cells, which changed shape as usual even when the cuticle was chemically disrupted, indicating that the cuticle is unlikely to play a guidance role. Extracellular matrices occupy a major portion of skeletal elements of animal body, and while the roles of extracellular matrices in cell signaling has been studied extensively, much less is known about how cells instruct shaping of extracellular environment, says Hayashi. Our study opened a new direction of research into morphogenesis of non-cellular parts of exoskeletons, which varies enormously in shape, color and function among insects. |
||||
|
||||
 |
| Copyright (C) CENTER FOR DEVELOPMENTAL BIOLOGY All rights reserved. |