| RIKEN Center for Developmental Biology (CDB) 2-2-3 Minatojima minamimachi, Chuo-ku, Kobe 650-0047, Japan |
December 14, 2011 A host of factors, both internal and external, are involved in determining whether a mitotic cell division will give rise to daughter cells of like or different types. Extrinsic signals can exert a variety of effects within the cell, from directly regulating the transcription of genes to remodeling the cytoskeleton. But how structural changes such as the latter translate to downstream effects on cell symmetry remains only poorly understood.
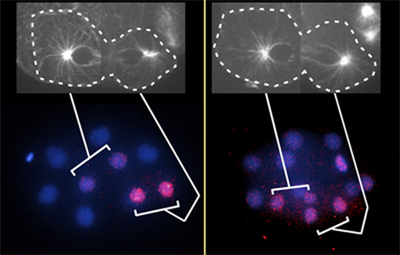
Recent work by Kenji Sugioka and others in the Lab for Cell Fate Decision (Hitoshi Sawa, Team Leader) has revealed a new mechanism by which external Wnt signaling generates mitotic spindle asymmetry, and thereby promotes asymmetric localization of ß-catenin within the nucleus. Published in Cell, this work reveals a novel means by which external signals can affect cellular properties in a cytoskeleton-mediated fashion in the roundworm C. elegans. Both Sawa and Sugioka have since moved to work at the National Institute of Genetics in Mishima, Japan. Early in roundworm development, a cell referred to as EMS receives external Wnt signals from a neighboring cell located posterior to it. Previous work has showed that this asymmetric signal causes WRM-1 (a C. elegans homolog of ß-catenin) to accumulate selectively at the anterior cortex of the EMS cell, which subsequently sets up an asymmetry during mitosis resulting in the accumulation of WRM-1 in the posterior daughter cells nucleus, and a second factor, POP-1, in the nucleus of the anterior daughter. The upshot of these allocations is that the anterior and posterior progeny take on different development fates, with the anterior daughter giving rise to muscle, and the posterior, gut. Suspecting that the cytoskeletal component known as microtubules might play a role in establishing this nuclear asymmetry, Sugioka treated EMS cells with a microtubule-disrupting chemical, and found that this indeed appeared to affect asymmetric WRM-1 and POP-1 localization in treated nuclei. Looking more closely at the behavior of microtubules during EMS cell division, the team found that astral microtubules increased in the cells anterior as mitosis progressed, with the greatest anterior-posterior differential occurring at the end stages of division. Inhibiting Wnt downstream factors WRM-1 and APR-1 by RNAi resulted in a decrease in anterior microtubules, indicating that their asymmetric localization is regulated by Wnt signaling. APR-1 co-localizes with WRM-1 at the anterior cell cortex, and it has been suggested that its mammalian counterpart (APC) has a stabilizing effect on microtubules. Sugioka used a GFP fusion protein to examine the distribution of APR-1 in dividing EMS cells. Early in mitosis there was no apparent asymmetry in its localization, but it gradually accumulated in the anterior side of the cell at later stages, strongly reminiscent of the changes in microtubule distribution observed the previous experiment. Inhibition of Wnt also reduced or eliminated this asymmetry, strongly suggesting that this signaling pathway controls the development of asymmetry by inducing unilateral localization of APR-1, leading to the asymmetric accumulation of microtubules. A subsequent experiment showed that APR-1 binds to and stabilizes the plus-ends of microtubules at the anterior cortex, suggesting a mechanistic basis for this effect. The team next used an optical (laser) disruption system to directly disrupt microtubule formation and observe the effects on the asymmetric nuclear localization of WRM-1 and POP-1. Perturbing anterior microtubule formation resulted in loss of asymmetric accumulation of these factors in nuclei, while dsirupting posterior microtubules conversely increased this asymmetry. Interestingly, when they used the same technique to induce microtubule asymmetry in Wnt mutants in which microtubules do not form asymmetrically, they found that the nuclear asymmetry of WRM-1 and POP-1 was restored. This set of findings makes a strong case for the role of microtubules in establishing the asymmetric nuclear accumulation of cell fate determinants in dividing C. elegans cells. It will be interesting to find out exactly what role microtubules are playing in conveying nuclear factors and establishing asymmetry, says Sawa. We will also be studying the question of how general this regulatory mechanism is to other biological contexts. | |||||
|
|||||
 |
| Copyright (C) CENTER FOR DEVELOPMENTAL BIOLOGY All rights reserved. |