| 独立行政法人 理化学研究所 神戸研究所 発生・再生科学総合研究センター |
| 2008年1月28日 |
哺乳類の受精卵は数回の卵割によって桑実胚と呼ばれる細胞塊をつくり、その後、内部が液体で満たされた胚盤胞と呼ばれる中空の球となる。胚盤胞は、外側を包む栄養外胚葉と内側に位置する内部細胞塊からなり、それぞれ将来、胎盤と胚本体を形成する。最近の研究で、この細胞分化の制御に1対の遺伝子Oct4とCdx2が重要な役割を果たすことが示されていた。Oct4とCdx2は互いに発現を抑制しあう転写因子で、Oct4を優位に発現すると内部細胞塊に、Cdx2を優位に発現すると栄養外胚葉に分化する。この発現バランスを調節するメカニズムについては諸説あるが、潜在的な上流因子の存在については未解明のままだった。 理研CDBの西岡則幸研究員と山元進司研究員(胚誘導研究チーム、佐々木洋チームリーダー)らは、マウスをモデルにした研究で、栄養外胚葉の分化にTead4と呼ばれる転写因子が必須であることを明らかにした。Tead4はCdx2の上流因子であることを示唆する結果が得られたという。この研究は、Mechanisms of Development誌に2007年11月17日付けでオンライン先行発表された。
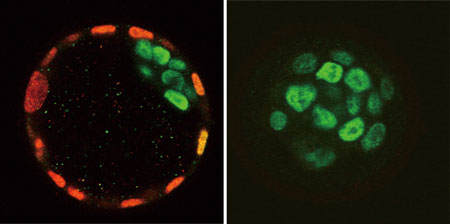
彼らは以前の研究で、Teadファミリー遺伝子が、着床後の発生を制御する正中線シグナルセンターの形成にかかわることを示していた。ところが今回、彼らがTead4を欠損する変異マウスを作成したところ、非常に興味深い結果が得られた。胚盤胞が胎盤に着床する前の、発生のごく初期に致死となったのだ。この思いがけない早死を受け、彼らは、着床前のマウス胚におけるTead4および関連遺伝子であるTead1、Tead2、Tead3の発現を再検証した。その結果、Tead1、Tead2、Tead4は4細胞期までに発現していることや、少なくともTead1とTead4は栄養外胚葉と内部細胞塊の両方で発現していることがわかった。 Tead4欠損マウスの胚発生を詳しく調べたところ、桑実胚期までは一見正常に発生したが、その後、細胞増殖は正常であるにもかかわらず、胞胚腔(胚盤胞内部の空洞)の形成は見られなかった。胞胚腔は、胚表面の隙間から液体が入り込んで内部を満たすことで形成されるため、細胞接着の異常によって阻害されることがある。しかし、Tead4欠損胚では、細胞接着や細胞の極性に異常はみられなかった。 次に彼らは栄養外胚葉に着目した。正常な胚盤胞の形成には、栄養外胚葉も極めて重要な役割を果たすからだ。栄養外胚葉の分化に必要とされるCdx2の発現を調べたところ、Tead4欠損マウスでは、最初の数回の卵割までは僅かに発現していたが、その後消失していることが明らかとなった。Cdx2依存的なFgfr2や、Eomesといった他の栄養外胚葉に特異的な遺伝子も発現していなかった。そこでOct4の発現を調べると、後期胚盤胞の栄養外胚葉では抑制されているはずが、胚全体に渡って発現していた。このことは、胚全体が内部細胞塊への運命を辿っていることを示唆していた。彼らは、Tead4欠損胚からES細胞を樹立し、これらの細胞がin vitroにおいて3胚葉全てに分化できることも示した。これは、胚本体の発生にTead4が必要でないことを示唆する結果だった。 佐々木チームリーダーは、「Tead4欠損胚で栄養外胚葉の形成が全く起きなかったことは、哺乳類胚発生における最初の細胞分化の機構を理解する上で、重要な手がかりになると考えています。しかし、胚の全ての細胞で発現しているTead4が、どのようにして一部の細胞においてのみ栄養外胚葉への分化を促進するのかは大きな問題であり、この仕組みを解明することが今後の課題です」と語る。 |
|||||
|
|||||
| Copyright (C) CENTER FOR DEVELOPMENTAL BIOLOGY All rights reserved. |