| 独立行政法人 理化学研究所 神戸研究所 発生・再生科学総合研究センター |
| 2008年7月14日 |
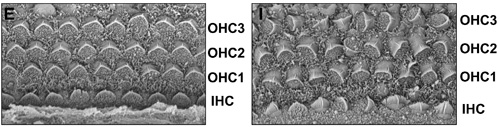
脊索動物の初期発生では中胚葉から脊索と呼ばれる構造が出来て体軸を規定する。胚発生ではこの脊索によって定められた体軸による方向性をもとに正しい位置に正しい組織・器官が形成され、この制御は胚発生において極めて重要な役割を持つ。今回山元進司研究員(胚誘導研究チーム、佐々木洋チームリーダー)は、この脊索に発現する遺伝子をスクリーニングし、脊索特異的な発現パターンを分子Cthrc1を同定した。さらに、このCthrc1が多様なWnt経路のうち平面内極性に関与するWnt/PCP経路を選択的に活性化し、内耳の有毛細胞の配向パターンを始めとする組織形成に関与していることを明らかにした。この研究成果はDevelopmental Cell誌7月号に発表された。 Wnt経路はC.elegansからヒトまで広く保存された分泌型のシグナル因子であり、胚発生において時間および位置特異的に発現し形態形成誘導因子,細胞極性決定因子,増殖分化調節因子など多様な機能を持つ。Wnt経路のうち最も良く知られているのはWntレセプターの一つであるFrizzledからβ-cateninを介して転写因子TCT/LEFを活性化する経路 (canonical Wnt pathway)であり、体軸形成などに関与する。この経路以外のnon-canonical Wnt pathwayは二種類存在し、それぞれWnt/Planar cell polarity(PCP:平面内極性、平面内の軸にそった極性)経路とWnt/Ca2+経路と呼ばれている。Wnt/Ca2+経路に関しては未だ不明の点が多いが、Wnt/PCP経路はRhoとその下流のROCK、及びRac1その下流のJNK活性化を通してアクチン重合を制御する。この経路は特に原腸形成や、内耳コルチ器にある有毛細胞の繊毛が正しい位置と方向性を持って並ぶために必要な経路であることが知られている。canonicalか、noncanonicalか、どちらのWnt経路を選択的に活性化する制御機構は未だ不明の部分が多かった。canonical Wntに属するWnt1、Wnt3a、Wnt8はFrizzledのコレセプターとしてLRP5、LRP6を用いシグナルを伝える。一方noncanonical Wntに属するWnt5a、Wnt11経路のコレセプターにはRor2がある。canonical Wntとnoncanonical Wntのどちらが活性化されるかはこれらのコレセプターの影響が大きい。 山元研究員らはマウス胚の発生過程において遺伝子スクリーニングを行った結果、脊索及び内耳において極めて特異的に発現する分子Cthrc1を同定した。Cthrc1は血管が障害されると発現上昇し細胞が移動しやすい状況を作るための分泌性の糖タンパク質であることが知られている。胚発生に伴う形態形成では分泌性シグナル分子を介した相互作用が細胞間のコミュニケーションの1つの方法として広く使用されている。山元研究員はCthrc1のマウス胚における脊索、内耳特異的な発現パターンに加え、分泌因子でもあることからこの分子に着目して研究を進めた。 Cthrc1ノックアウトマウスは特に異常を示さなかったが、山元研究員はその特徴的な発現パターンからWnt/PCP経路に関与する可能性を考え解析を進めた。その結果、同じく平面内極性に関与する遺伝子Vangl2のヘテロ変異体とCthrc1ノックアウトマウスを掛け合わせると、中脳領域で神経管閉鎖不全がおこるなど平面内極性に支障を来している場合の特徴的な表現型を示した。また、同じく平面内極性が深く関与する内耳蝸牛管内にあるコルチ器の有毛細胞も、細胞自体の形成には問題ないものの、本来あるべき方向性を持った配毛パターンが見られなかった。
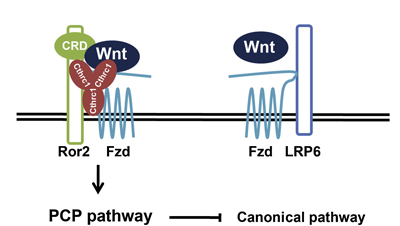
更に詳しい解析を進めた結果Cthrc1はnoncanonical Wnt経路のコレセプターであるRor2と結合してWnt/PCP経路を活性化し、組織の平面内極性に関与することが分かった。Cthrc1はRor2のみならずWnt及びFrizzledにも結合することから、Wnt-Frizzled-Ror2間の結合性を促進し、Wnt/PCP経路を活性化していると考えられる。また、Cthrc1はcanonical Wnt経路のコレセプターであるLRP5、6とは結合しないことから、noncanonical Wnt経路のみを選択的に活性化することが示唆された。 Wnt経路はコレセプターや下流の因子のバリエーションにより様々な場面で多様な機能を持つ。しかしWntがいかにしてコレセプターを選びそのバリエーションを生むのかは分かっていなかった。今回の研究で山元研究員らはCthrc1がRor2に結合しWnt/PCP経路を選択的に活性化することを解明し、Wntによる多様な機能が如何に制御されているのか、その機構の一つが明らかになった。
一方でCthrc1のノックアウトマウスが目立った異常を見せないことから、佐々木チームリーダーは「私たちは、Cthrc1の機能を補完する遺伝子がほかにも存在するのではないかと考えており、そのような因子を同定し、その機能を解析してゆくことが、多様な機能を持つWntの作用の制御機構を理解するには
必要だろうと考えています。」と語る。今後Wntシグナル経路が時間的、空間的に如何に制御されているのか明らかになることが期待される。
|
|||||||
|
|||||||
 |
| Copyright (C) CENTER FOR DEVELOPMENTAL BIOLOGY All rights reserved. |