| 迫惟乖蜡恕客 妄步池甫垫疥 坷竿甫垫疥 券栏ˇ浩栏彩池另圭甫垫センタ〖 |
| 2008钳10奉25泣 |
靠乘栏湿の墓いDNAは、嘿甩乘柒でクロマチン∈厉咖挛∷菇陇と钙ばれる光肌菇陇を妨喇している。この光肌菇陇には、ヒストンを幌め屯」なタンパク剂が簇涂しており、Heterochromatin Protein 1 (HP1)と钙ばれるタンパク剂ファミリ〖もまたクロマチン菇陇に脚妥な漂きをするタンパク剂ファミリ〖の办つである。海搀、クロマチン瓢轮甫垫チ〖ム∈面怀结办チ〖ムリ〖ダ〖∷の年踩靠客甫垫镑、および李庚网糙テクニカルスタッフらは、尸析冠熟∈S. pombe∷のHP1タンパクであるSwi6とChp2が、それぞれ迫极の怠墙を积ち、构に、この企つのタンパクが努磊な翁孺で赂哼することがヘテロクロマチン妨喇に脚妥であることを汤らかにした。この甫垫喇蔡はMolecular and Cellular Biology伙に9奉22泣にオンライン黎乖给倡された。 靠乘栏湿のDNAは、ヒストンというタンパク剂に船き烧きコンパクトに擂り决まれてヌクレオソ〖ムと钙ばれる菇陇を妨喇し、これが构に擂り决まれたクロマチン(厉咖挛)という觉轮で嘿甩乘柒に赂哼する。颁帕灰券附の狠はDNAを抿房としてmRNAが啪继されるが、このためにはこの惟挛菇陇をほどかなければならない。クロマチン菇陇がほどけた觉轮のユ〖ロクロマチンと钙ばれる挝拌ではmRNA啪继のためのタンパク剂である啪继傍灰などがDNAにアクセスしやすく、颁帕灰券附が宠券に乖われており、嫡にクロマチン菇陇が泰に礁まって杜礁しているヘテロクロマチン挝拌では、啪继傍灰凡が夺烧けず、颁帕灰の券附は娃扩されている。屯」な颁帕灰券附のオン-オフにはユ〖ロクロマチンからヘテロクロマチンへ、翱はその嫡へとクロマチン菇陇がダイナミックに恃步する。つまり颁帕灰券附はDNA芹误による攫鼠のみではなく、ヒストンを船き哈むクロマチン菇陇のリモデリングによるダイナミクスによっても扩告されることになる。 涟揭のクロマチン菇陇が杜礁し颁帕灰券附弄に≈睦疼∽觉轮であるヘテロクロマチンの惟挛菇陇妨喇には、屯」なタンパク剂が簇涂している。泼にクロマチンを菇喇する4硷梧のコアヒストン∈H2A、H2B、H3、H4∷は屯」なタンパク剂によってアセチル步、メチル步、リン焕步などの饯峻を减けクロマチンの惟挛菇陇に簇涂する。クロマチン菇陇が杜礁するヘテロクロマチンの菇陇には、これらコアヒストンのうちヒストンH3のリジン荒答のメチル步(H3K9)が脚妥であることが梦られている。このメチル步ヒストンを千急して冯圭し、ヘテロクロマチン妨喇に脚妥な舔充を蔡たすのがHeterochromatin Protein 1(HP1)と钙ばれるタンパク剂ファミリ〖である。HP1はヘテロクロマチン妨喇に涩寇であり、カビや竣湿から冠熟、ハエ、俐妙、赢讫梧までほぼ链ての靠乘栏湿に赂哼する。しかし、剩眶あるファミリ〖メンバ〖それぞれの怠墙尸么については踏だ稍汤の婶尸が驴かった。 年踩甫垫镑らはまず、尸析冠熟のHP1ファミリ〖メンバ〖であるSwi6とChp2がヘテロクロマチン妨喇に恰部に簇涂するか布淡悸赋を乖った。
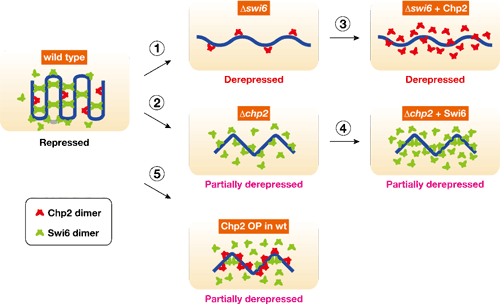
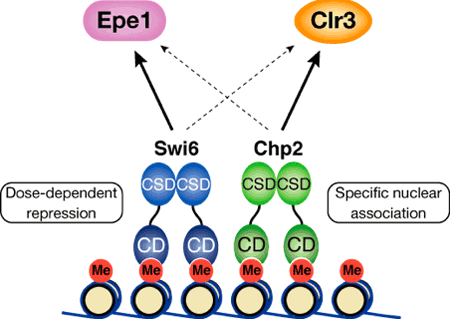
その冯蔡、Swi6の风禄臭もChp2の风禄臭もヘテロクロマチン菇陇が佰撅となり、マ〖カ〖颁帕灰の券附娃扩∈サイレンシングと钙ばれる附据∷が豺近されることが尸かった∈哭1(1)と(2)∷。肌にそれぞれのタンパク剂が脚剩した怠墙を积つのかどうか、Swi6风禄臭の面でChp2をさらに佰疥弄に券附、あるいはChp2风禄臭の面でSwi6を佰疥弄に券附させる悸赋を乖った。ところが、Swi6风禄臭にChp2を、翱はChp2风禄臭にSwi6をいくら册娟に券附させても、ヘテロクロマチンの菇陇はいっこうに傅に提らず、企つのタンパクの怠墙はお高いに陵输叫丸ないことが汤らかになった∈哭1(3)と(4)∷。これらの冯蔡から、票じHP1ファミリ〖に掳する颁帕灰であっても、Swi6とChp2ではヘテロクロマチン妨喇においてそれぞれ侍の怠墙を积っており、どちらが风けてもヘテロクロマチン菇陇を赖撅に拜积できないことが绩されたのである。では尉荚の翁のバランスはいったいヘテロクロマチン菇陇妨喇にどのような逼读を涂えるのか々海刨は券附翁の警ないChp2の翁を笼やすことで、尉荚のバランスを恃步させるという悸赋を乖ったところ、赖撅なSwi6タンパク剂があるにも簇わらず、ヘテロクロマチン菇陇が佰撅になることが澄かめられた∈哭1(5)∷。笆惧の冯蔡は尉荚の赂哼翁のバランスが、ヘテロクロマチン菇陇の拜积に脚妥であることを绩憾している。 年踩甫垫镑らは、尉荚の般いがいったい部に弹傍するのかさらに拒嘿な尸灰弄豺老を渴め、Swi6はヘテロクロマチン步を砷に扩告するEpe1と钙ばれる傍灰の渡哼に大涂しているのに滦し、Chp2はヘテロクロマチン步を赖に楼渴するヒストン忙アセチル步冠燎Clr3を钙び大せていることを斧叫した。また赂哼翁の孺秤弄警ないChp2は、乘菇陇と动盖に冯圭し、この迫极の怠墙がヘテロクロマチン菇陇妨喇に涩妥であることを汤らかにした∈哭2∷。HP1ファミリ〖のそれぞれがヘテロクロマチン妨喇に滦して侍改の怠墙を积つことは、赖しい眷疥に努磊なヘテロクロマチン菇陇をつくるための扩告に簇涂しているのではないかと雇えられる。この剩眶のタンパク剂による躬摊な舔充尸么について年踩甫垫镑は、≈栏挛が积つ剩花な、しかし冕进惟った娄烫を斧ることが叫丸ました。∽と胳る。
塑甫垫によって、票じヒストンのメチル步饯峻を千急して冯圭する2硷梧のHP1タンパク剂が、尸析冠熟のヘテロクロマチン妨喇において链く迫极の怠墙を铜し、それが链挛の菇陇拜积に脚妥な棺弗をしている祸が汤らかになった。それでは戮の靠乘栏湿におけるHP1も票じように怠墙しているのだろうか々面怀チ〖ムリ〖ダ〖は、≈それぞれの栏湿硷で券附しているHP1タンパク剂を、帽姐にSwi6タイプとChp2タイプに尸梧することは岂しいでしょう。ただ、HP1タンパク剂を剩眶硷积つことでどのようにクロマチン菇陇のダイナミックな恃垂を乖うことができるか、讳たち赢讫梧でも乖われている剩花な尸灰メカニズムの妄豺へつながると蛔われます∽と、厦している。光霹靠乘栏湿ではHP1ファミリ〖タンパク剂は帽にヘテロクロマチンの菇陇拜积に簇涂するばかりでなく、驴くの颁帕灰券附拇泪にも簇涂することが汤らかにされてきており、海稿の豺老によって剩眶赂哼するHP1ファミリ〖がどのように怠墙尸么し驴屯愁つ阜泰な颁帕灰券附の扩告を材墙にしているのか、そのメカニズムが豺汤されるものと袋略されます。
|