
ヒトを含む多細胞動物の体は多数の細胞が社会をつくり組織・器官を形成している。この多細胞体制の形成・維持には細胞同士の接着と認識機構が必須であり、この機構に中心的な役割を果たすのがカドヘリンである。カドヘリンは細胞外から細胞質側に細胞膜を貫通して存在し、この細胞外に出ている部分が隣り合う細胞のカドヘリンに結合し、細胞同士を接着する。この時カドヘリンは、細胞質側でカテニンを始めとするタンパク質群と複合体を形成し、更に細胞骨格タンパク質を動員した複雑な装置を形成することにより、細胞の形態や移動をも制御している。この細胞間接着装置が破綻すれば、発生過程における組織の形成は支障をきたし、また、がん細胞などで見られる異常な浸潤・転移へとつながる。この複雑な接着装置の解析は動物の発生現象や細胞のがん化を理解する上で非常に重要である。
今回、高次構造形成研究グループ(竹市雅俊グループディレクター)孟文翔研究員らは、この複雑なタンパク質複合体である細胞間接着装置が上皮細胞の接着帯においてどのように構成されているのかを解明する過程で、この装置が細胞骨格タンパク質である微小管と相互作用して細胞の形態維持に関与していることを明らかにした。この研究成果はCell誌11月28日号に掲載された。
カドヘリンが形成する細胞接着構造である接着結合(adherens junction:AJ)のうち、極性をもつ上皮に特徴的な接着結合を接着帯(zonula adherens:ZA)と呼ぶ。接着帯は上皮細胞の頂端部側に存在して細胞周囲を取り巻いており、細胞間隙に存在するE-カドヘリンから、細胞内の骨格タンパク質へとつながる複合体が細胞間接着装置を形成する(図1)。
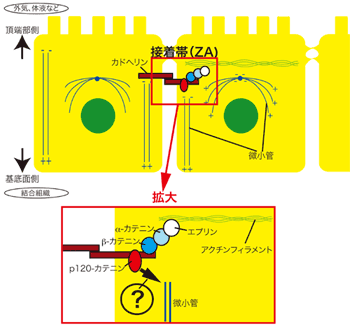 |
図1 上皮細胞の接着帯(ZA)
上皮細胞には外気や液体にさらされる頂端部側と結合組織に接着する基底面側という極性を持つ。頂端側では細胞と細胞が接着した接着帯(ZA)が存在し、その接着帯にはカドヘリンを含む細胞間接着装置が存在する。
|
|
この接着装置で最も良く知られるのは、カドヘリンに細胞質側で結合するβ-カテニンから、α-カテニンなどを介して細胞骨格タンパク質であるアクチンフィラメントへとつながる部分である(図1接着帯拡大図)。近年、同じく細胞骨格タンパク質である微小管もまた細胞間接着と深く関与しているらしいことが解ってきた。しかし、微小管が細胞接着に関与する詳細なメカニズムについては不明の点が多かった。
高次構造研究グループは以前、カドヘリンの細胞質ドメインに結合するp120-カテニン(以下p120)が微小管と相互作用して細胞移動などに関与することを明らかにしていた。孟研究員らはこのp120が微小管のダイナミクスを如何に制御するか(図1(?)部分)更なる解析を進めた。まず、p120がどのようなタンパク質と相互作用するのか解析を行ったところ、PLEKHA7というタンパク質がp120結合タンパクとして同定された(図2)。
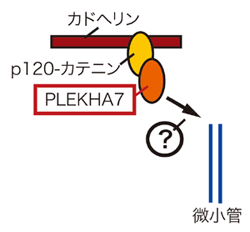 |
図2 p120結合タンパクPLEKHA7の同定
PLEKHA7はp120依存的に細胞間接着部位、特に接着帯に存在し、p120及びカドヘリンの接着帯への局在を制御する。PLEKHA7がないとE-カドヘリン、p120は接着帯に局在出来ない。
|
|
その他の実験から、PLEKHA7はp120に結合して接着帯に局在し、カドヘリン、p120を含む細胞間接着装置を構成するタンパク群の統合に必要であることが示唆された。
次にこのp120-PLEKHA7複合体と微小管の相互作用(図2(?)部分)を明らかにするため、孟研究員らはPLEKHA7に結合するタンパク質のスクリーニングを行った。その結果、新規タンパク質であるNezhaを同定した(図3)。
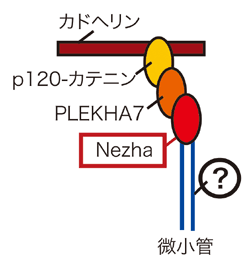 |
図3 PLEKHA7結合タンパクNezhaの同定
NezhaはPLEKHA7依存性に接着帯に局在する。NezhaがないとPLEKHA7、E-カドヘリン、p120の接着帯における局在パターンが乱れる。
|
|
更にこのNezhaが微小管と如何なる関係にあるか解析したところ、Nezhaは微小管のマイナス端に局在することが分かった(図4-1)。多くの微小管はマイナス端が中心体と結合し、細胞の様々な領域にプラス端をのばしている(中心体微小管)。しかし興味深いことに、Nezhaが結合する微小管のマイナス端は接着帯に局在し、これをアンカー点としてプラス端をのばしていた。これらの結果よりNezhaが結合する微小管は中心体からのびる微小管ネットワークとは別の“非中心対微小管”であることが解った(図4-2)
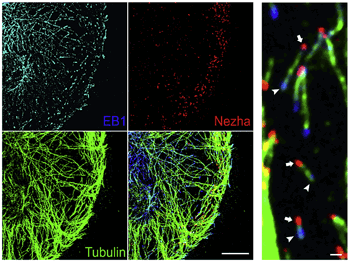 |
図4-1 Nezhaの微小管マイナス端への局在
Nezha(赤)は、微小管(緑)のプラス端に局在するEB1(水色)と対極のマイナス端に存在することが解る。この微小管は中心体とは結合しない“非中心対微小管”である。
|
|
|
図4-2(Movie) Nezhaと微小管のダイナミクス
Nezha(赤点)のある微小管マイナス端から緑色のシグナル(微小管プラス端に局在するタンパク質EB1)が延びていくように移動するのが解る。Nezhaの名前の由来は,西遊記の登場人物で、武器を投げる様子がこのムービーとよく似ていることから命名された。
|
|
これらの結果より、Nezhaと微小管との深い関与が予測されたことから、孟研究員らはPLEKHA7、Nezhaを含む複合体が微小管の機能に如何に関与するのか(図3(?)部分)解析を行った。微小管は細胞内のレールとしての役割を持ち、その上をモータータンパク質が移動している。孟研究員らはこのモータータンパク質のうち、微小管プラス端からマイナス端に移動するKIFC3に着目し、PLEKHA7及びNezhaとの関係を解析した。その結果、KIFC3は微小管を使って接着帯に局在しており、NezhaまたはPLEKHA7がないとKIFC3は接着帯に局在出来なかった。一方KFC3が存在しないとE-カドヘリンが正しくZAに分布しないことも解った。これらのことからKIFC3はPLEKHA7とNezhaに依存して接着帯に局在し、カドヘリンを含む細胞間接着装置の維持に関与していると結論された(図5)。しかし、モータータンパク質であるKIFC3が何を接着帯まで運ぶのかは解っておらず、今後の解析が待たれる。
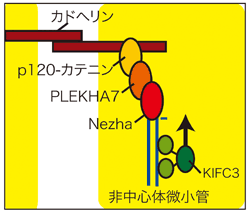 |
図5 接着帯におけるE-カドヘリン、p120、PLEKHA7、Nezhaを含む細胞間接着装置
|
|
KIFC3、PLEKHA7、Nezhaが失われると、上皮細胞は頂端部側の構造を保てず、細胞の形態が扁平になる。これらのタンパクは協調して細胞間接着装置の機能に関与すると考えられる。
細胞形態や細胞間接着の安定性は、がん細胞の異常な増殖、浸潤に深く関与する。孟研究員は「このPLEKHA7、Nezhaによる制御機構が生体内で発生やがん細胞の増殖・浸潤などに如何に関与するか解明したい。」と語る。また、今回新規に同定されたタンパクNezhaの、非中心体微小管に特異的に結合するという特徴は非常に興味深い。非中心体微小管の動態や機能は不明の部分が多く、Neza-非中心体微小管が果たす細胞生物学的役割について、様々な角度からの研究が進行中である。
|
| Copyright (C) CENTER FOR DEVELOPMENTAL BIOLOGY All rights reserved. |
|
