| 独立行政法人理化学研究所 発生・再生科学総合研究センター |
2013年12月5日 |
細胞が特定の化学物質(誘引物質)の方向に移動する性質は走化性と呼ばれ、血球細胞や神経細胞などで免疫応答や組織形成に重要な働きを担っている。単細胞真核生物である細胞性粘菌は飢餓期において、走化性を用いて集合し多細胞体を作る。走化性の分子メカニズムはほ乳類の白血球細胞と良く似ており、わずか数十マイクロメートルの細胞体の前後でほんの数パーセントの濃度差を感じ取って走化性を示す。一方、細胞は周囲に勾配が無い状態でも細胞内部のゆらぎによりランダムに方向転換しながら自発的に動くことが知られている。そのようななか、一体どのようにして細胞は誘因物質の微小な濃度差を内部環境のゆらぎを乗り越えて検知し、増幅して、正しい方向に動くことができるのだろうか。 理研CDBの柴田達夫ユニットリーダー(フィジカルバイオロジー研究ユニット)らは細胞性粘菌を使った研究で、細胞内部の自発的な極性形成機構の詳細を蛍光イメージングの統計解析によって明らかにし、そのメカニズムを数理モデルによって再構築することに成功した(Journal of Cell Science 誌 2012年11月号)。さらにその自己組織化する極性機構が、誘因物質への高い感受性と頑強性を助ける仕組みについて、このモデルを用いて明らかにした。この研究は理研QBiCと共同で行われ、Biophysical Journal 誌の2013年11月号に掲載された。(以降、自己組織化する極性機構とする)
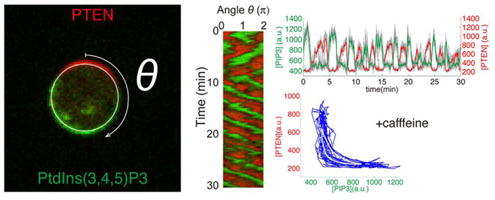
細胞性粘菌は誘因物質cAMPの濃度勾配に応答して細胞膜上でイノシトールリン脂質の代謝反応を引き起こす。高いcAMP濃度に接した領域ではPI3キナーゼによってホスファチジルイノシトール4,5-二リン酸(PIP2)からホスファチジルイノシトール3,4,5-三リン酸(PIP3)が産生され、逆に低い領域では脱リン酸化酵素であるPTENによってPIP3からリン酸基がとれてPIP2が生成される。そしてPIP3濃度の高い領域ではアクチン繊維が重合して仮足が伸び、走化性を引き起こす。 柴田らは2012年の論文において、まず実際に粘菌を使い、誘因物質の濃度勾配がない状態で、かつ細胞自身が誘引物質のcAMPを産生することを阻害して細胞間の相互作用を排除した上でPIP3タンパク質の動態を見た。すると、細胞膜上でPIP3の発現ドメインがランダムに現れ、それが細胞膜上を動く様子が見られ、内部の自発的な極性形成機構が確認された。PTENはPIP3とは互いに排他的に局在していた。また細胞膜上のあるポイントでの濃度の増減について解析し、統計処理を行った。すると両者の関係は単純な酵素と基質の相互作用のみでは表すことが出来ないことが示唆され、これを詳細に解析するとPIP3が少ない場所によりPTENが集まりやすいという新たな知見が得られた。次に反応拡散モデル(空間内のある物質の濃度について、その化学反応と拡散反応を考慮に入れた変化を表す数理モデル)を用いてシミュレーションを行うと、上の観察結果と一致しPIP3ドメインが持続的に現れるパターンが得られた。さらにパラメーターの値を調節すると、ドメインが現れたり消えたりを繰り返すパターンが得られ、これはcAMP産生を阻害しない場合の観察結果と一致していた。以上より与えたモデルが適正であることが分かる。数理モデルの解析から細胞の自発的な極性形成、すなわち自己組織化する極性機構は神経等でよく見られる興奮系の性質(少しの刺激でも、しきい値を越えれば十分な応答を作り出す性質)を持つことがわかった。
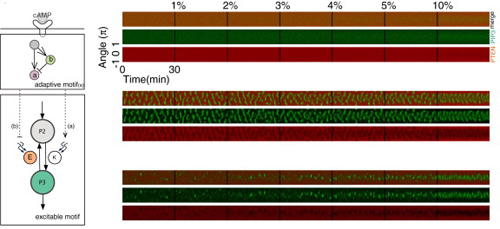
次の論文では、誘因物質の勾配を入れた場合について解析した。まず実際に細胞性粘菌にcAMPを加えたところ、あらかじめ持続的ドメイン形成パターンがあった場合と、一過的ドメイン形成パターンがあった場合の両方においてcAMPの濃度が高い方向にPIP3ドメインの分布が偏ることを確認した。次に確率的な反応拡散モデルを構築し、2パターンに加え、細胞内部の自己組織化する極性機構が働いていない静止状態を作り出し、それぞれcAMP濃度を段階的に上げてシミュレーションを行った。作用させるcAMPの濃度が上がると、あらかじめ細胞内部に持続的、または一過的ドメイン形成パターンがあった場合の方が、静止状態に比べてより小さなcAMP勾配でも強い応答が見られた。このことから自己組織化する極性機構はcAMP勾配への応答性を促進することが示唆された。 また、どのような機構によって濃度勾配の方向に沿ったドメイン形成の偏りが実現されているのかを検証するために、細胞膜上のPIP3ドメインの形成場所と頻度を定量的に解析した。すると誘因物質の勾配がある場合には無い場合よりドメインの形成頻度は増え、濃度勾配が上がると勾配の方向に対するドメイン形成位置の正確性が上がることが分かった。ドメインの持続時間やPIP3の濃度は勾配とは相関が見られなかった。以上のことから、濃度勾配の傾斜が増加するとともに、ドメイン形成の頻度と方向の正確性が上昇して、細胞は高い感受性と頑強性を持った走化性応答を実現できることがわかった。これは、外部環境に応じて細胞内のドメインの形成されやすさに極性が生じ、濃度が高い場所ではドメイン形成のしきい値が下がって形成しやすくなることを示していた。 細胞性粘菌はcAMPの進行する波に向かって走化性を示すことで集合し、多細胞体を作る。ここで正しく波の来る方向を感知するには濃度が増えて行く方向(波が向かってくる側)のみに動き、減って行く方向(波をおいかける側)へは動かない必要がある。この認識機構について、柴田らは上流に適応反応(刺激に対して一過的に応答し、その後元に戻る反応。視覚や嗅覚などの感覚細胞にしばしば見られる性質)、下流に自己組織化する極性機構を組み合わせた数理モデルを検討した。このモデルに、空間的にcAMPの濃度の進行する波を適用したところ、濃度が上昇するタイミングにのみ細胞は応答した。この結果より、細胞は濃度が上がっているのか、下がっているのかということ自体を認識して自己組織化する極性機構のしきい値をコントロールし、波の向かって来る方向に動いていることが示唆された。 柴田ユニットリーダーは「今回、これまで知られている分子生物学の知識と、蛍光イメージングのデータの詳細な検討から、実験を比較的よく説明する数理モデルを構築できました。その結果から、分子機構や、勾配感知の振る舞いについて、新たな知見が得られ、今後実験的な検証が待たれます。また、自己組織化を用いた機能発現については、同じ概念が他の多くの場合にも適用されるはずです。」と語る。
|
|||||||||||||||||
 |
| Copyright (C) CENTER FOR DEVELOPMENTAL BIOLOGY All rights reserved. |