| RIKEN Center for Developmental Biology (CDB) 2-2-3 Minatojima minamimachi, Chuo-ku, Kobe 650-0047, Japan |
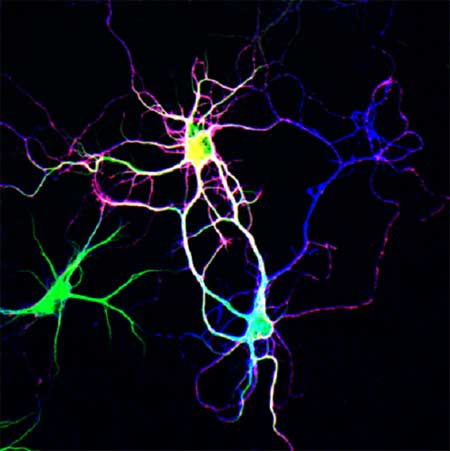
The Togashi study started by examining the behavior of two members of the nectin family of adhesion molecules. Previous reports had shown that nectin-1 (N1) localizes mainly to the presynaptic neuronal membrane, while nectin-3 (N3) prefers the postsynaptic side of the junction. Nectins tend to bind heterophilically, meaning that they adhere more strongly to nectins of different type, and have been shown to serve as recruiters for cadherin-family adhesion proteins, which have also been implicated in the regulation of synapse formation. This set of preliminary evidence prompted the group to take a closer look at nectin activity at the synaptic scale. Turning first to localization, Togashi used immunostaining to show that, in hippocampal neurons, N3 is distributed about equally in both axons and dendrites, while N1 is far more abundant in axons. When neurites of different type encountered each other and began to from synapses, both N1 and N3 became concentrated at the point of contact, but interestingly, when a pair of dendrites happened to cross paths, no such concentration was evident. With these initial observations pointing to possibly differential roles for the two nectins in neurites, the Takeichi group next tested the effects of their overexpression. Neurons transfected with N1 showed marked increases in the dendritic concentration of this nectin as well as abnormalities in the behavior of the neurites as they extended outward from the neuronal somata. Normally, axons and dendrites extend radially from the neuronal cell body, but in the N1 transfected cells, axons became entwined around dendrites and dendrites occasionally looped back upon themselves, creating a briar-like entanglement. In a converse experiment using cells in which the N1 gene was knocked out, dendritic spines failed to develop stable contacts with axons in cultures of mutant hippocampal neurons, despite the presence of other genetic markers of synaptogenesis. As transmembrane proteins, the nectins comprise both extracellular and intracellular (or cytoplasmic) domains. Experiments utilizing partial constructs indicated that the N1 cytoplasmic domain is responsible for its axonal preference. Togashi tried a mix-and-match approach to test for possible effects on neurite affinities and patterning, and found that when the N1 extracellular domain was yoked to the N3 cytoplasmic moiety, the resultant phenotype was the same as that for the simple overexpression of N1.The reverse pairing produced a less severe phenotype, suggesting that the N1 extracellular domain is responsible for the molecule’s regulatory effect on neurite association. The case for interaction between the two nectins in promoting neuritic associations was further strengthened by experiments in which N1- and N3-transfected cells were cultured together; as might be expected, their dendrites became tightly intertwined. Additional tests in which neurons were introduced onto a mosaic layer of transfected and untransfected HEK293 cells revealed that when a dendrite developed a stable contact with an N1-transfected cell, N1 molecules from the cell became intensely concentrated at the dendritic points of contact. This was not true for axons, which did not appear to recruit nectins of either type. Nectins do not make their way to the sites of synapse formation unaccompanied; immunostaining showed that β-catenin, a component of the cadherin cell adhesion machinery, was also present at nectin binding sites, and that its concentration increased proportionally to the intensity of nectin accumulation. The overexpression of either type of nectin also produced an increase in levels of N-cadherin, irrespective their neuritic associations. And, while N-cadherin overexpression had no discernible effect on neurite patterning, neurons in which αN-catenin was deleted (resulting in impairment of cadherin function) were unresponsive to the dendritic abnormalities associated with nectin-1 transfection. This thoroughgoing investigation led Togashi et al. to propose a new model of the nectins’ roles in neurite interaction. In this model, N1 molecules on the axon preferentially bind their N3 counterparts on dendrites, which has the secondary effect of promoting homophilic cadherin binding. Without N1, cadherin-based adhesion occurs much less intensely, resulting in weaker connections between cells, while in N1-transfected cells, opportunities for binding increase, causing dendrites (which are normally N1-free) to associate with other dendrites, and even normal axodendritic contacts to overstabilize. “We had previously thought that cadherins alone were sufficient for the initial contacts between axons and dendrites, but we now realize that, by cooperating with Ig-superfamily molecules, the cadherins can be utilized by neurons in more sophisticated ways.” says Takeichi. “This sort of interaction between cadherins and Ig-superfamily molecules may be involved in other cell-cell recognition systems as well, a possibility that we are looking forward to studying in the future.” |
|||||||||
|
|||||||||
 |
|||||||||
| Copyright (C) CENTER FOR DEVELOPMENTAL BIOLOGY All rights reserved. |