| Sox2 sits atop different transcriptional networks in ESC and TSC self-renewal |
 |
 |
Nov 7, 2013 The first unmistakable differentiation events in mammalian development take place just as the blastocyst is forming, with the outer layer of cells committing to form the extra-embryonic trophectoderm, and the inner cell mass giving rise to the earliest embryonic lineages. When grown in culture, these inner cell mass cells are known as embryonic stem cells (ESCs), and show the ability, known as pluripotency, to give rise to every embryonic lineage, as well as the ability to proliferate indefinitely by self-renewal in vitro. Trophectoderm likewise arises from trophectoderm stem cells (TSCs), which form the embryonic component of the placenta. The genetic regulation of such stemness in ESC, TSC and neural stem cells relies in part on the function of the transcription factor Sox2, but how the function of this protein functions differs across these cell types has never been resolved.
A new in-depth study by Kenjiro Adachi and colleagues in the Lab for Pluripotent Stem Cell Studies (Hitoshi Niwa, Project Leader) reveals that Sox2 sits atop two different networks of stemness genes in mouse ESCs and TSCs. Conducted in collaboration with the CDB labs for Genomic Reprogramming, Functional Genomics, Systems Biology and Genome Resources and Analysis, this work was published in Molecular Cell.
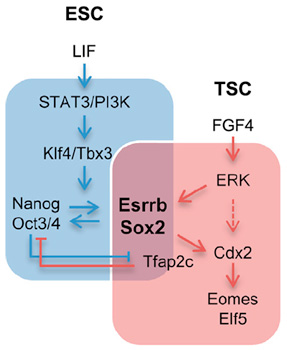 |
A schematic of genetic networks regulating stemness in early embryonic cells.
In mouse ESCs, Sox2 is controlled by LIF, and in TSCs by FGF4. The switch of Sox2s binding
partner from Oct4 to Tfap2c changes the set of downstream genetic interactions, inducing
a transition from ESC to TSC state. In this scheme, Oct4 and Tfap2c are mutually inhibitory. |
Previous work by the Niwa lab had shown that deletion of Oct4, a known pluripotency gene, caused ESCs to transform into TSCs, which are known to rely on the activity of FGF4. In the present study, Adachi and colleagues examined changes in gene expression during this transition, and determined that in TSCs, FGF4 activates Sox2 and a second gene, Esrrb. They found that by misexpressing these genes in TSCs, they could maintain their stemness even in the absence of FGF4. In contrast, in mouse ESCs, Sox2 is under the control of LIF, indicating to the team that Sox2 might interact with different sets of genes to maintain stemness and inhibit differentiation in trophoblast and embryonic stem cells.
What then is responsible for the switch in Sox2 targets? Adachi et al. next looked at Sox2 DNA and protein binding, and discovered something interesting: Sox2 does not act alone, but functions by binding with other transcription factors in a cell context-dependent fashion. While it binds Oct4 in ESCs, in TSCs it partners with Tfap2c; these changes in binding partner trigger conformational changes that enable interactions with independent networks of target genes in these stem cell types.
The idea that combinations of the activity of various signaling pathways helps to give rise to cellular diversity is widely accepted, but it is generally thought that the factors involved and the order of activation within a pathway are fairly uniform, says Niwa. But what we have shown here is that changes in upstream and downstream factors can play significant roles in driving cell diversity, and we suspect perhaps evolutionary processes as well.
|
