細胞は、1つが2つに分裂することで増えていく。体を構成する体細胞では、まず、核のなかにある染色体が2倍に複製され、半分ずつに分離する。同時に細胞質も分かれ、1セットずつ染色体をもつ娘細胞が2個つくられる。このとき、細胞内の物質が均等に分配された上に、細胞外の環境にもまったく違いがなければ、2つの娘細胞はまったく同一のものになり、異なる細胞が生み出される余地はない。
「19世紀には、受精卵の卵割を観察することで、もとの細胞とは異なる娘細胞ができることが分かっていた」。非対称細胞分裂研究グループの松崎文雄博士は、そう話す。観察の対象にされたのは、ホヤやウニなどの海産動物の受精卵だ。受精卵は卵割を繰り返すことで細胞数を増やしていくが、8細胞くらいになるまでは、分裂のようすが外からもよくみえる。例えば、ホヤの場合、最初の卵割では細胞質が均等に分配されることが分かる。しかし、次の卵割では黄色くみえる何らかの物質が均等に分配されないことが観察されていた。「研究者たちは、この黄色い物質の分配のされ方が、将来どのような細胞になるかといった運命決定に影響することを認識していた」と松崎博士。
異なる娘細胞がつくられることは、大きさや形の違いからも確認されていた。ツノガイでは、最初の卵割で一方の娘細胞のみに小さなこぶができ、そのこぶは、次に分裂した2個の娘細胞のどちらかだけに受け継がれる。発生学者のウィルソン博士は、このこぶを除去すると2個の細胞が同一のものになってしまうことを実験で確かめ、非対称に分配されるこぶに含まれる因子が細胞の運命を決定することを示した。
このように細胞質が均等に分配されない分裂は「非対称分裂」と呼ばれる。複数種の細胞からなる多細胞生物ではかならず非対称分裂が起きるが、単細胞生物でも非対称分裂を起こすものが知られている。例えば、出芽酵母は、親酵母から芽のような娘細胞が出芽して、それが大きくなることで増えるが、親酵母の性(メイティングタイプ)と娘酵母の性はかならず異なる。つまり、性にかかわる因子が非対称に分配されていることになる。出芽酵母は、普段はこのような非対称分裂を繰り返すことで増えるが、厳しい生存環境になると分裂をやめて胞子をつくる。豊かな環境になると、異なる性の胞子どうしが合体して、再び非対称分裂を行うようになる。
このように、非対称分裂は多くの生物でみられるが、発生のどの時期に始まるかは、さまざまだ。ツノガイの場合は最初の卵割から非対称だが、マウスでは少なくとも4細胞までは全く均等に分割しているといわれている。ただし、一見して色や形、大きさが同じ細胞でも、将来異なる細胞に分化することもあり、その場合には非対称分裂の範疇に入る。
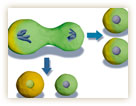
非対称分裂を起こす要因はなんなのか。大きく分けると二つある。一つは、細胞中の物質の分布に偏りがある場合だ。もう一つは、細胞の外から与えられるシグナルに偏りがある場合。細胞は、こうした物質やシグナルの偏りに応答して、自らの「前後や左右」といった極性を獲得していく(図1)。
ただし、細胞内の物質の偏りによって細胞極性が得られても、極性の方向と分裂の方向が一致しなければ、同じ極性をもつ同一の細胞が2個できるだけなので、非対称分裂にはならない。極性方向と分裂軸を一致させることが、非対称分裂の必須条件となるのだ(図2)。

1960年代以降、分子生物学者のブレンナー博士は、透明な体をもつ線虫を用いて非対称分裂のしくみを詳細に調べた。体長1ミリほどの線虫は、その名のとおり細長いひものような生物で、神経や咽頭、腸、筋肉など、動物の基本的な構造を備える。ブレンナー博士は、線虫の発生過程を生きたまま観察し、一つの受精卵が成虫にいたる過程を詳細に追跡した。その結果、雌雄同体の成虫は正確に959個の体細胞からなり、例えば、そのうち302個が神経細胞になるなど、一つひとつの細胞がたどるべき系譜を明らかにした。
この細胞系譜によると、線虫の場合、受精後最初の分裂から非対称だが、その極性は精子の侵入位置によって決められていた。2回目の分裂以降は、親細胞がほかの細胞とどの部位で接しているか、といった情報によって非対称分裂が起き、生じた娘細胞はそれぞれの運命決定に従って、神経や消化管などの細胞に分化していくことが分かった。
その後、非対称分裂に異常をもつ線虫の変異体が多く見つかり、受精卵に不均等に分配して細胞極性をつくるpar遺伝子群などが同定されている。線虫は、非対称分裂を遺伝子、分子レベルで研究する上で非常に重要な役割を果たしたのだ。
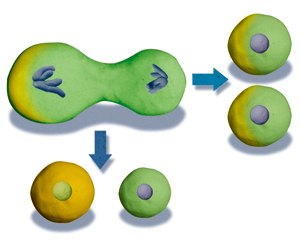
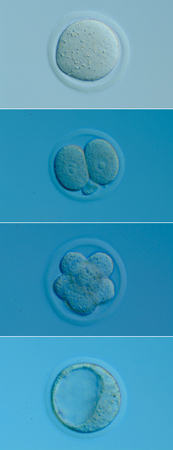
発生初期の卵割において、まったく均等に分裂している場合には、そのうちの一つの細胞を取り出して別の子宮に移植すると、あたかも一つの受精卵のように振る舞って個体へと成長する。発生におけるこうした能力を「全能性」という。
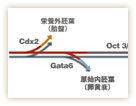
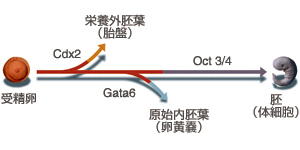
マウスの場合、4細胞に至る時期までは、全能性を保っているとされている。しかし、8細胞期から16細胞期にかけて、全能性は失われてしまう。この時期になると、外側の細胞と内側の細胞が生じ、それぞれの細胞が異なる運命を獲得するからだ。その結果、外側の細胞は将来胎盤をつくる栄養外胚葉になり、内側の細胞は体をつくる内部細胞塊へと分化する(図3)。内部細胞塊は体を構成する全ての細胞を生み出せるが、もはや栄養外胚葉になることはできない。そのため、内部細胞塊は「多能性」幹細胞と呼ばれ、それだけを子宮に移植しても発生が進むことはない。細胞はこのようにして分化の第一歩を踏み出すことになる。
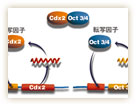
多能性幹細胞研究チームの丹羽仁史博士は、細胞分化の第一段階に起きる現象を、遺伝子のレベルで明らかにしようとしている。「とくに、細胞がどのようにして多能性を獲得し、それを維持するのかを明らかにしたい」と話す。2000年には、内部細胞塊を培養したES細胞が多能性を維持するために重要なOct3/4遺伝子を同定。その後、2005年には栄養外胚葉を分化誘導するCdx2遺伝子の存在を明らかにしている。
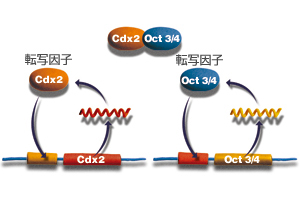
そこで丹羽博士は、Oct3/4遺伝子とCdx2遺伝子がどのように相互作用しているのか、マウスの受精卵をつかって検証してみた。すると、それぞれの遺伝子からつくられる産物が、それぞれの遺伝子を自己活性化させる機構をもつことが分かった。つまり、Oct3/4分子がOct3/4遺伝子の転写を、Cdx2分子がCdx2遺伝子の転写を活性化する機能をもっていたのだ。また、Oct3/4分子とCdx2分子が共存すると、互いに結合して、転写活性化能を失うことも明らかとなった(図4)。つまり、両方の分子が存在する場合には、少しでも量が多い方の遺伝子発現のみが誘導されるらしい。
これら2つの遺伝子の初期発生における発現について調べると、「最初は全ての細胞で両方の遺伝子が発現しているが、10~16細胞の桑実胚期になると、内側の細胞ではOct3/4遺伝子のみが、外側の細胞ではCdx2遺伝子のみが発現するようになる」と丹羽博士は説明する(図5)。「内側の細胞では、Oct3/4の発現量がしだいにCdx2の発現量を上回るようになり、最終的にはCdx2は発現しなくなる。結果として、内側の細胞は栄養外胚葉になることから逃れ、多能性を維持したまま内部細胞塊を形成する。外側の細胞では、これとまったく逆のことが起こるのだろう」。
それでは、これら2つの遺伝子の発現のバランスは、何をきっかけに崩れるのだろうか。そこに細胞極性のメカニズムがあると丹羽博士は予測している。「内側と外側の細胞ができるこの時期には、何らかの因子の不均等な分配があり、Oct3/4およびCdx2の発現に影響を与えているのではないか。今後は、そのような極性をつくっている分子の同定も目指したい」と語る。
丹羽博士の研究チームは2002年に、将来卵黄嚢になる原始内胚葉を分化誘導するGata6遺伝子も同定している。「これら3つの遺伝子で、哺乳類の初期の発生における細胞分化は、おおよそ説明できると思う」と丹羽博士。「Oct3/4は哺乳類のみにみられる遺伝子で、発生初期にしか発現しない。つまり、この遺伝子がCdx2やGata6をあやつることにより、胎盤や卵黄嚢を獲得させつつ、多能性幹細胞を維持するシステムをつくり上げていったのではないだろうか(図6)」とコメントし、Oct3/4の進化的起源についても探っていきたいという。
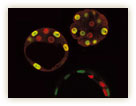
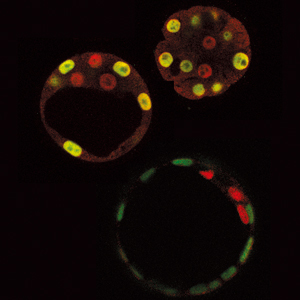
非対称分裂による細胞分化が見られるのは、発生の最初期だけではない。むしろ、細胞が多様化するほどに、非対称分裂は重要になってくる。1995年、神経幹細胞を経て神経細胞が分化するメカニズムを研究していた松崎博士は、偶然にも非対称分裂に行き着くことになる。「ショウジョウバエの突然変異体のなかから、神経発生に異常がみられるものを探しだし、その原因遺伝子としてプロスペロという遺伝子を同定した。しかし奇妙なことに、プロスペロは神経幹細胞で発現しているにもかかわらず、分裂後にできた一方の娘細胞でしか働いていなかったのです」。松崎博士は、当時をそう振り返る。
プロスペロの発現のようすを詳細に調べたところ、神経幹細胞においてプロスペロタンパク質は確かに発現しているものの、その分布には明らかな偏りがみられた(図7)。「神経幹細胞が分裂期に入ると、プロスペロは細胞表層の特定部分に移動していた。その結果、一方の娘細胞のみに受け継がれ、神経細胞やグリア細胞などへの分化を誘導するようだ」と松崎博士。プロスペロタンパク質はDNAに結合して遺伝子の発現を調節する転写因子だ。そのため、機能を果たすには細胞核のなかに入る必要がある。プロスペロは娘細胞に受け継がれて初めて核内に移行する、という松崎博士の研究結果も、これと整合性がとれていた。
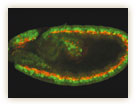
神経幹細胞は、上皮細胞がある決まった方向に落ち込んで形成される(図8)。松崎博士は、上皮細胞に由来するほかの幹細胞についても解析を行い、プロスペロを不等に分配する幹細胞の極性が、上皮細胞の表裏の極性に基づいていることを見出した。最近になって、いくつかの研究グループが、プロスペロタンパク質の不等分配をつくりだす分子機構を明らかにしつつあり、上皮細胞の極性との関連が証明されつつある。
「プロスペロ遺伝子は哺乳類にもあるが、非対称分裂を制御しているという証拠はまだない」と話す松崎博士。今後は、マウスでも実験を行い、哺乳類だけにみられる非対称分裂のメカニズムと、多くの生物種が共通してもつ非対称分裂のメカニズムに迫りたいとしている。
細胞分化と多能性維持の機構に迫る丹羽博士の研究は、応用分野においても期待を集めている。内部細胞塊に由来するES細胞は、多能性を維持しているため、人為的にさまざまな細胞をつくることができる。そのため、創薬研究や移植医療に利用できると期待されているのだ。「多能性の分子機構が完全に解明できれば、ES細胞を均一な状態のまま増殖させたり、さらには、分化した体細胞を初期化し、多能性を獲得させることも可能かもしれない」と丹羽博士は語る。
高度な非対称分裂システムは、一方で多能性を獲得させ、一方で細胞を多種多様に分化させる機構をもたらした。松崎博士は「非対称分裂による多様化のメリットは、細胞ごとに異なる機能を果たし、役割分担を行うことで、より高度な生命システムを築けることだろう」と話し、そのメカニズムの解明にさらなる意欲をみせる。
38億年におよぶ生命史において、約10億年前に現れたとされる多細胞生物は、非対称分裂のしくみを利用して爆発的な進化を遂げてきた。生命多様化の鍵ともいえる非対称細胞分裂。その核心が解き明かされる日はそう遠くないのかもしれない。

