
CDBからのニュース、お知らせを掲載しています。
体には頭尾軸、背腹軸、そして左右軸の3つの軸がある。私たちの体は外見的には左右対称のようで、心臓や肝臓、腸などの形状・配置は左右非対称。複雑な臓器を体の中にコンパクトに収納する上で、この非対称性は不可欠だ。左右軸は発生の初期に、頭尾軸と背腹軸の決定を受けて最後に定まるが、哺乳類の左右軸決定機構は実にユニークだ。胎児期初期に「ノード」と呼ばれる領域が一過的に形成されるが、ここの細胞は各々1本ずつ繊毛を持っている。それらがくるくると回転することで右から左への一方向の体液の流れ(ノード流)が生じ、その刺激を受けて体の左側を規定するシグナル因子NodalやLeftyが誘導されて、左右対称性が破られ左右軸が形成されるのだ。しかしノード細胞が頭尾・背腹軸の位置情報をきちんと把握し、それらを踏まえて正確に左右軸を決定する仕組みの詳細は、いまだ未解明な点も多い。
理研CDBの峰岸かつら研究員(個体パターニング研究チーム、濱田博司チームリーダー)らは、マウス初期胚を用いて、一方向のノード流が生み出されるに至る分子メカニズムを詳細に解析。Wnt5が頭尾軸に沿ってグラデーション状に発現することで細胞内平面極性シグナル経路が刺激され、ノード細胞にある繊毛の付け根が尾側に移動することで、一方向性のノード流を生み出す仕組みを明らかにした。本成果は科学誌Developmental Cellに2017年3月13日付で掲載された。
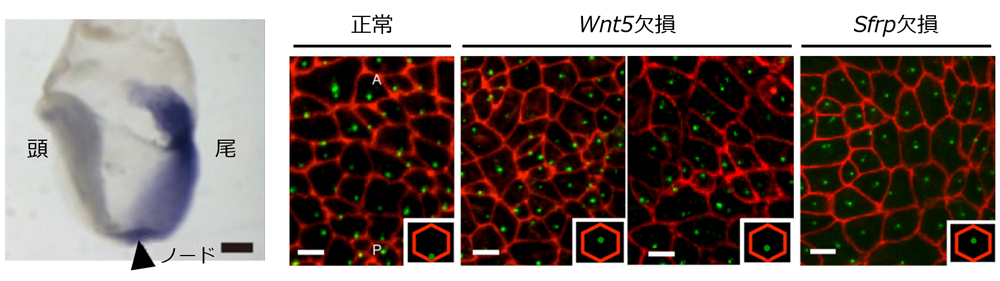
ノードは初期胚の遠位部に現れるお椀型のくぼみ構造で、マウスでは胎生8日前後の約12時間という限られた期間に観察される。200~300個の細胞が並び、各々が持つ1本の繊毛を時計回りに回転させている。繊毛の付け根に存在する基底小体は、当初はノード細胞の中央に位置しているが、発生の進行と共に徐々に尾側に移動し、それに伴って繊毛の回転軸も尾側に傾いていく。これによって、個々に小さな渦を作っていただけの繊毛の回転が、ノード全体として右から左へ向かう大きな一方向の流れを生み出すようになる。研究チームは過去の研究で、この物理的な「流れ」こそがNodal・Leftyの発現部位の決定に重要な要因であることを示した。また、これまでの報告から、基底小体の尾側への移動には平面内細胞極性(planar cell polarity:PCP)シグナルが関与していることが示唆されていたが、先に形成された頭尾軸の情報がどのようにして基底小体の移動の向きに結び付けられるのか、その分子機構は不明だった。
そこで峰岸らは、ノード出現時期の胚全体において、頭尾軸に沿ってグラデ―ション状に発現しているWnt5に着目。Wnt5は頭尾軸の形成に寄与することが知られ、頭側で弱く、尾側で強く発現している。Wnt5のノックアウト(KO)マウスを詳細に解析すると、基底小体は中央から動かなかった。また、Wntの拮抗因子であるSfrpはWnt5とは逆向きのグラデーション状に発現するが、このSfrpのKOマウスも同様に基底小体が尾側に移動しなかった。さらに、Wnt5やSfrpを過剰に発現させてグラデーションを壊しても、基底小体は移動しなかった。これらのことから、Wnt5とSfrpのグラデーション状の発現がノード細胞の基底小体を尾側に移動させるのに重要であることが分かった。SfrpはWntと逆向きのグラデーションを作ってWntを抑制することで、Wntのグラデーションをより明瞭にしていると考えられる。
では、Wntシグナルのグラデーションをノード細胞はどのように感知するのだろうか。峰岸らは次に、個々の細胞内で極性を生み出す常套手段であるPCPシグナルに着目し、既知の頭側因子Vanglと尾側因子Dvlの発現を確認。すると、確かにWnt5のグラデーション状の発現の後、各細胞の頭側または尾側の細胞膜付近で特異的に発現していた。さらに今回、PCPシグナル因子の一つPrickleもまた頭側因子として機能することを新たに同定。このKOマウス胚では、基底小体は細胞ごとにばらばらの方向に移動し、一定方向のノード流ができなかった。さらにノードにおいてWnt5を細胞ごとにモザイク状に欠損させると、基底小体は正常細胞/Wnt5欠損細胞によらず、様々な方向に移動した。尾側への移動の成否は細胞ごとにランダムだが、移動の向きは近隣の細胞集団で同じ方向に向かっているようだった。このことから、Wntシグナルのグラデーションの感知には、隣接する細胞同士の細胞間コミュニケーションが寄与している可能性が示唆された。
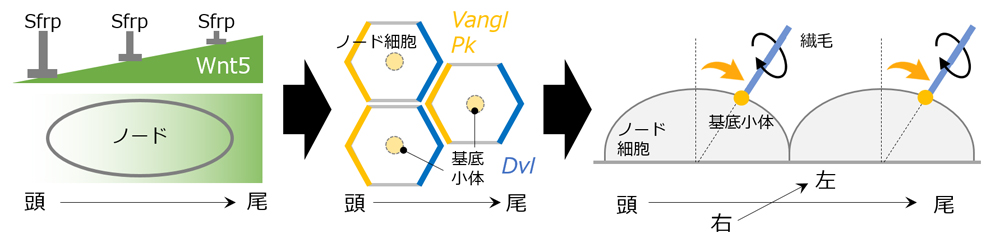
「Wnt5の発現量の差を見分けてPCPシグナルは活性化しますが、このとき、ノード細胞同士の細胞間コミュニケーションによって情報を補正していると考えられます。ノード細胞はWntシグナルが規定した頭尾軸を正確に捉え、それに直行する左右軸を生み出すことに成功している。細胞内のシグナルと周辺細胞からの情報を統合する、巧妙なメカニズムがそこにはあるはずです」と濱田チームリーダーは語る。「今回の研究では、基底小体の移動のイニシャルキュー(最初のきっかけ)をWntシグナルが担っていることを示しました。しかし、ノード細胞は単一細胞内のWnt濃度のわずかな差をどのように正確に感知しているのか?細胞間コミュニケーションを担う分子は?基底小体を移動させる仕組みは?など、謎は尽きません。今後も左右の対称性が破られる仕組みに迫りたい。」
| 掲載された論文 |
|---|
![CDB 理化学研究所 発生・再生科学総合研究センター [RIKEN CENTER FOR DEVELOPMENTAL BIOLOGY]](http://www.cdb.riken.jp/wp-content/themes/cdb/images/common/fLogo2.png)
〒650-0047 兵庫県神戸市中央区港島南町2-2-3
TEL : 078-306-0111
FAX : 078-306-0101
E-mail : cdb[at]cdb.riken.jp [at]を@に変えてメールしてください。

