
News and Announcements from the CDB
The inner ear is located in the inside of the head, however at the earliest stages of embryonic development, it forms as a thickening of the ectoderm (epithelia) on the outer surface of the embryo, referred to as the otic placode. This placode invaginates into the mesenchyme of the future head region, eventually budding off to form an otic vesicle, the precursor of inner ear organs. Invagination of the otic placode, similar to many epithelial invagination processes, involves two phases: first, there is an expansion of the basal membrane of otic cells, followed by a constriction of the apical side of otic cells. Whereas the molecular mechanisms that drive basal membrane expansion in the first phase of otic invagination are beginning to be understood, the mechanisms controlling constriction of the apical side in the second phase remain relatively unknown.
Now, new work published in Developmental Biology by research scientist Xiaorei Sai of the Laboratory for Sensory Development (Raj K. Ladher, Team Leader) and colleagues has uncovered a molecular pathway coordinating the apical constriction of otic cells during the second phase of otic invagination. Using chicken embryos as a model, they demonstrate that localized activation of Ras homolog gene family member A (RhoA) protein at the apical cell junctions triggers a signaling pathway leading to recruitment and activation of non-muscle myosin-II, an actin-binding protein, to initiate apical constriction of the otic placode.
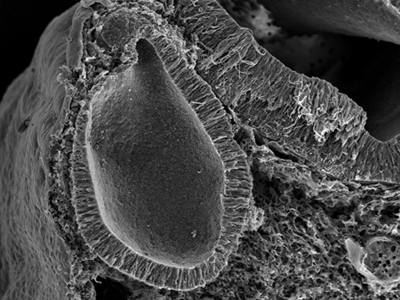
Scanning electron micrograph showing the inner ear after 50 hours of development.
The inner ear, which formed on the surface ectoderm, has now completed its invagination,
and is embedded within the head mesenchyme
A previous study from this laboratory (Sai and Ladher, 2008) showed that basal activation of non-muscle myosin-II instigated basal expansion of the otic placode in phase 1 of otic invagination; myosin-II is activated by basally localized FGF signaling, which leads to depletion of actin filaments on the basal side, and consequently, basal expansion. Studies of similar invagination processes, such as gastrulation and lens placode formation, by other groups have indicated that activation of myosin-II is also important for apical constriction, where activated myosin-II triggers a signaling cascade leading to constriction of actin filaments on the apical side. Thus, these findings suggest that activation of myosin-II at apical and basal sides of the otic placode involve different molecular cues, resulting in contrasting activities for myosin-II on actin filaments.
So what are the molecular cues at work at the apical side? Sai et al. focused their sights on the RhoA protein, a member of the Rho family of small GTPases, which is known to regulate actin cytoskeleton dynamics and has also been implicated in promoting apical constriction of epithelial cells during invagination processes in other model systems. In these systems, RhoA triggers Rho-associated coiled-coil protein kinases (ROCKs) to phosphorylate myosin light chain (MLC) to activate myosin-II, leading to constriction of actin filaments. In the case of the otic placode, Sai et al. observed a rise in RhoA expression during phase 2 of otic invagination, with high levels detected in the apical junction complex. RhoA activation assays on otic placode extracted from embryos during this same period also confirmed the presence RhoA activity. When RhoA activity was knocked down using small RNAi, they detected reduced levels of active apical MLC as well as actin in the apical junctions, and a larger apical surface area than control embryos suggested that minimal or no constriction had taken place. Thus, these results highlight the importance of RhoA for apical constriction. They also looked into whether ROCKs, potential RhoA downstream factors, also played a role in phase 2 constriction by inhibiting ROCK expression and found that otic invagination was blocked in addition to disruption of the localization of actin and activated MLC to apical junctions, indicating ROCKs were also needed for apical constriction.
The team next attempted to determine the molecule triggering RhoA activity. Rho family small GTPases are generally switched on by guanine exchange factors (GEFs), so they performed a screening for apically localized GEFs with using antibodies. The ArhGEF11 protein was deemed a putative RhoA GEF since its expression became concentrated in the apical area as otic placode invagination progressed, overlapping with expression of junctional marker ZO-1. Knocking down normal ArhGEF11 activity levels led to reduced activity of myosin-II and localization of actin to apical junctions. ArhGEF11 expression was not affected when RhoA was knocked down, thereby indicating that ArhGEF11 was upstream of RhoA and also involved in apical constriction.
Studies on neural tube formation had reported that a planar cell polarity signal, Celsr1, was needed to activate myosin-II through RhoA and its upstream GEF. Thus, the team speculated that Celsr1 may be an upstream regulator of RhoA and its GEF, ArhGEF11, to enhance apical constriction in the otic placode. Expression experiments showed that while weak Celsr1 expression is detected in the ectoderm just as the otic placode is beginning established, during the phase I of otic morphogenesis, its expression is upregulated and localized to the apical junction, reaching a peak during the phase 2. When Celsr1 expression levels were reduced in otic placodes during the phase 2, they found reduced activity and localization of actomyosin at the apical cell junction, resembling what was seen in ArhGEF11 knockdown experiments.
Based on their data, they proposed a model for apical constriction seen in phase 2 of otic invagination, where the planar cell polarity protein Celsr1 is upregulated in the apical otic placode as otic morphogenesis begins, and accumulates at the apical cell junctions. Celsr1 then recruits ArhGEF11 to the apical junction, which in turn activates RhoA, a key factor needed to trigger ROCKs to activate myosin-II, leading to constriction of actin filaments anchored to the apical junction.
“It is clear that epithelial remodeling in the neural tube and during inner ear invagination share a common morphogenetic toolbox,” says Ladher. “Thus findings in one of these processes could be used and adapted to understand conditions with clinical significance, such as neural tube defects, body wall closure defects and wound healing.“
| Link to article | |
|---|---|
| Related link |
How cells go pear-shaped(June 23, 2008) |
![CDB [RIKEN CENTER FOR DEVELOPMENTAL BIOLOGY]](http://www.cdb.riken.jp/en/wp-content/themes/cdb_en/images/common/fLogo2.png)
2-2-3 Minatojima-minamimachi, Chuou-ku, Kobe 650-0047, Japan
TEL : +81-78-306-0111
FAX : +81-78-306-0101
E-mail : cdb[at]cdb.riken.jp

